

表达PINK1蛋白的胶质母细胞瘤患者生存结局更好(上)
发布时间:2026-05-09 10:28:01 | 阅读:次| 关键词:表达PINK1蛋白的胶质母细胞瘤患者生存结局更好(上)
- [案例]常规产检发现胎儿脑血管畸形,孩子会不会很危险,又该
- [案例]周五开启与巴教授面对面 | 脑干胶质瘤疯长125%,早期手术
- [案例]这个位置手术易瘫痪?18岁少年高位颈髓手术顺利完成
- [案例]中脑顶盖肿瘤“懒癌启示录“——切还是不切,这是个问
- [案例]开颅也看不见的脑瘤如何应对?国际专家运用“可视化技
- [案例]室管膜瘤全切术后3年未复发,距离痊愈还有多远?
James T. Rutka(鲁特卡)教授曾任世界神经外科学院院长,30多年来深耕儿童神经外科研究,带领团队不断取得突破,从脑瘤分子分型、精准治疗到新药治疗与微创治疗,为全球神经外科疑难病患儿带来了新的希望。Rutka教授团队发表了研究《PINK1 Is a Negative Regulator of Growth and the Warburg Effect in Glioblastoma》(PINK1是胶质母细胞瘤生长和Warburg效应的负调控因子),本文为该研究的简述,重点介绍其研究方法与设计。
摘要
增殖中的癌细胞具有一系列特征性代谢改变,包括高糖酵解率、高乳酸生成量以及线粒体代谢的重编程,这些改变为肿瘤细胞(包括胶质母细胞瘤)的持续增殖提供了关键代谢产物。然而,这些生物过程同时会产生氧化应激,必须通过及时清除活性氧来维持细胞内平衡。
研究团队利用无偏倚逆转录病毒功能缺失筛选技术,在正常转化的人星形胶质细胞中证实,线粒体PTEN诱导激酶1(PINK1)是Warburg效应的调节因子,同时也是胶质母细胞瘤生长的负调控因子。研究结果显示,PINK1的缺失通过ROS依赖性的缺氧诱导因子-1α稳定化,以及丙酮酸激酶肌肉同工酶2(PKM2)活性降低(这两者均为有氧糖酵解的关键调节因子),来促进Warburg效应。
从作用机制来看,PINK1通过调控FOXO3a(氧化应激和超氧化物歧化酶2的主要调节因子)来抑制ROS水平和肿瘤生长。这些发现凸显了PINK1与ROS平衡在的正常细胞和肿瘤细胞中均具有举足轻重的地位。
在大量人脑肿瘤标本(包括胶质母细胞瘤,样本量n > 900)中均观察到PINK1的表达缺失,且与患者的不良生存预后密切相关。在体内实验中,PINK1过表达能够显著减弱原位小鼠异种移植模型和果蝇转基因胶质母细胞瘤模型中的肿瘤生长。
引言
胶质母细胞瘤是所有胶质瘤中最为常见且致死性最高的亚型,患者平均生存期约为12-16个月。通过受体酪氨酸激酶激活的Ras信号通路激活以及p53缺失,是驱动人类疾病和小鼠胶质母细胞瘤模型中肿瘤形成的两个关键分子改变。癌症基因组图谱(TCGA)计划进一步验证了这些通路,同时也发现了此前在胶质母细胞瘤中未被识别的新改变。然而,这些改变中的绝大多数很可能是"乘客"改变,即并不直接参与癌症发生发展的改变。功能基因组学策略通常被用于识别驱动癌症发生、进展和治疗耐药的关键基因,这最终是对大尺度测序策略的重要补充。基因插入策略(包括Sleeping Beauty转座子方法)的实施,已经帮助研究人员鉴定出若干新的癌症相关基因。研究团队假设,在相关胶质母细胞瘤通路(包括Ras或p53信号)中存在改变的非转化星形胶质细胞,可以通过随机逆转录病毒插入来灭活参与肿瘤起始和/或进展的基因,从而实现转化。
通过逆转录病毒筛选,研究团队鉴定出PTEN诱导激酶1(PINK1)是肿瘤细胞依赖的众多细胞过程的负调控因子。PINK1是一种线粒体丝氨酸/苏氨酸激酶,在家族性帕金森病患者中存在突变,并可调节从线粒体自噬到活性氧产生和氧化磷酸化等多种生物学功能。众所周知,肿瘤抑制基因和癌基因的功能失调经常会影响到细胞内的ROS水平。癌细胞持续面临平衡氧化应激和控制ROS水平的挑战,因为ROS同时发挥着促生长和抗生长的双重作用。本研究报告,PINK1通过调节线粒体氧化磷酸化、有氧糖酵解和ROS水平来抑制胶质母细胞瘤的生长。
材料与方法
细胞培养:从p53-/-(1周龄)、NMU-P0(CD1小鼠)建立和表征原代星形胶质细胞培养物。胎儿人星形胶质细胞购自Lonza公司。U87和T98G细胞系购自ATCC。SF188细胞系由Chris Jones博士(英国伦敦癌症研究所)惠赠。所有细胞系均已于2011年通过短串联重复序列(STR)分型进行了表征和鉴定。细胞系在添加10%胎牛血清的DMEM培养基中生长。胶质瘤干细胞的培养方法如先前所述。
耗氧量和糖酵解通量测定:使用Seahorse XF96分析仪测量耗氧量(OCR)。细胞在其常规生长培养基中培养,并用无缓冲培养基重悬后接种于XF96培养板中。在转移至XF96分析仪之前,将细胞置于无CO2的37°C培养箱中,在无缓冲培养基中平衡90分钟。依次加入1.2 μmol/L寡霉素、1.0 μmol/L FCCP和1.0 μmol/L鱼藤酮,测量基础OCR。通过Seahorse XF96分析仪测量细胞外酸化率(ECAR),作为糖酵解通量的指标。
蛋白质印迹分析和免疫组化:蛋白质印迹分析和免疫组化实验均按先前所述方法进行。所用一抗及其稀释度如下:β-肌动蛋白、PINK1、半胱氨酸次磺酸、HIF1A、PKM2、纽蛋白、V5、PDHK1、磷酸化PDHE1α、PDHE1α、HK2、LDHA、磷酸化FOXO3a Ser294、FOXO3a、HA-Tag、SOD2。免疫组化用一抗:PINK1(1:100)和Ki67(1:100)。
体内小鼠模型实验:所有小鼠均按照病童医院(SickKids)的动物护理规范饲养,实验方案经审查批准后实施。在NOD-SCID小鼠中进行立体定向引导的颅内注射。通过在裸鼠NOD-SCID小鼠侧腹注射与Matrigel混合的2×10⁶个细胞进行侧腹成瘤实验。定期监测小鼠肿瘤生长情况,并用卡尺测量肿瘤体积。
果蝇研究:所有实验果蝇杂交均在25°C的昼夜节律培养箱中进行,用于产卵和幼虫发育。在游走期晚三龄幼虫阶段解剖果蝇幼虫脑,用于肿瘤体积比较和免疫组化分析。所用一抗包括:小鼠抗Repo和兔抗磷酸化组蛋白H3。荧光成像和体积测量方法如先前所述。
其他方法:溴脱氧尿苷(BrdU)细胞增殖实验、caspase活性实验、克隆形成实验、己糖激酶(HK)活性和PKM活性实验均如先前所述进行。
统计分析:所有实验均重复三次,适当时报告均值和标准误。多组间比较采用ANOVA分析,随后进行事后Dunnett检验(与一个对照组比较)或事后Tukey检验(识别各亚组间差异)。两组间直接比较使用非配对双尾Student t检验。生存分析使用对数秩(log-rank)检验。显著性阈值设定为*P < 0.05;**P < 0.01;***P < 0.001。
结果(其三)
研究最终共发现9个重要结果,受篇幅所限,本文先展示其中三个,其余结果将在后续文章中呈现,敬请关注。
逆转录病毒基因捕获及捕获克隆的功能表征
研究团队从正常CD1-ICR小鼠、新生超活化Hras小鼠和p53缺失小鼠建立了原代星形胶质细胞培养物。为了鉴定导致早期传代星形胶质细胞转化的功能缺失事件,研究人员用含有剪接受体(紧随荧光素酶报告基因和IRES嘌呤霉素抗性标记上游)的逆转录病毒基因捕获载体转导星形胶质细胞。通过嘌呤霉素筛选,共获得32个抗性克隆,其中14个表达荧光素酶。在这14个背景各异的表达荧光素酶的基因捕获克隆中,有9个能够在软琼脂糖中生长(这是衡量细胞转化能力的经典指标)。尽管NMU处理产生了5个嘌呤霉素抗性的表达荧光素酶克隆,但NMU并未介导星形胶质细胞转化。反向PCR技术鉴定了9个锚定非依赖性基因捕获克隆中7个的确切基因插入位点。其中GT-15和GT-16此前已在胶质瘤生物学中有过报道。本研究新鉴定的四个基因此前未涉及胶质母细胞瘤发病机制,研究人员对其细胞生长、抗凋亡和侵袭能力进行了进一步表征。
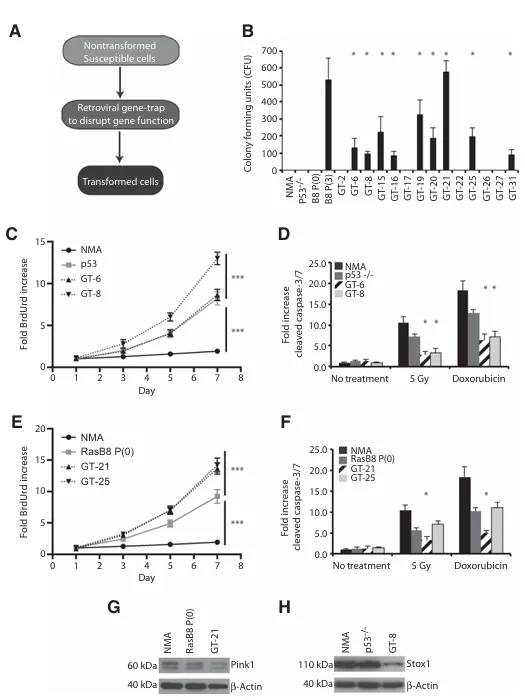
逆转录病毒基因捕捉的总体策略及捕获克隆的功能特性分析。(A)逆转录病毒基因捕捉的总体策略示意图。(B)表达萤火虫荧光素酶的捕获克隆在软琼脂培养试验中表现出显著的生长优势(9/14),这与未转化的星形胶质细胞形成鲜明对比。(C)基于p53背景的基因捕捉克隆的BrdUrd增殖试验结果。(D)针对基因捕获克隆的激活型caspase-3和-7 ELISA检测结果。(E)基于RasB8背景的基因捕捉克隆的BrdUrd增殖试验。(F)对来自RasB8星形胶质细胞的基因捕获克隆进行了激活的caspase-3和-7 ELISA检测。(G-H)Western印迹分析显示,与亲代细胞相比,捕获的克隆中Pink1和Stox1的表达水平有所降低。上方的条带代表完整长度的Pink1,下方的条带则代表Pink1在整合入线粒体后所形成的成熟形式。*P < 0.05;***P < 0.001
在p53-/-背景下,研究者在以下基因中鉴定出插入事件:GT-6和GT-8。与亲本p53-/-细胞相比,通过7天的BrdUrd增殖实验测量,GT-8细胞(而非GT-6细胞)的增殖能力显著增强。与亲本p53-/-细胞和未修饰的小鼠星形胶质细胞相比,GT-6和GT-8在暴露于5 Gy辐射或1 μmol/L阿霉素后均表现出caspase活性降低(即抗凋亡能力增强)。与对照细胞相比,GT-6和GT-8在用5 Gy辐射处理时集落形成数量增加。
在Ras-B8 P0背景下,研究者表征了GT-21和GT-25两个克隆。与对照相比,通过7天的BrdUrd掺入测量,GT-21和GT-25细胞均表现出增殖增加。与对照细胞相比,GT-21和GT-25在暴露于5 Gy辐射和阿霉素处理后显示caspase活性降低。与对照细胞相比,GT-21在用5 Gy辐射处理时集落形成增加,并且GT-21细胞是唯一表现出侵袭性增加的细胞。综上所述,GT-21在所有四个测试实验中均表现出显著改变。蛋白质印迹分析进一步证实,这些基因捕获事件改变了两个顶级候选基因PINK1和STOX1的蛋白质表达水平。
PINK1缺失改变正常人星形胶质细胞代谢
为了深入研究顶级候选基因PINK1在正常人类细胞系统中对细胞生长、氧化应激和线粒体功能的作用,研究团队在正常胎儿人星形胶质细胞和多个已建立的人胶质母细胞瘤细胞系中进行了功能验证实验。在原代人胎儿星形胶质细胞培养物中可检测到PINK1表达,但在大多数胶质母细胞瘤细胞系和外植体培养物中呈低表达或阴性表达。细胞分级分离分析结果表明,PINK1主要定位于线粒体。由于PINK1缺失对正常人类星形胶质细胞的影响尚不清楚,研究人员通过两个靶向PINK1的混合shRNA构建体,以及一个将V5表位标记的PINK1重新导入敲低细胞的PINK1拯救细胞系,生成了稳定的PINK1敲低克隆。与乱序对照和PINK1拯救细胞相比,PINK1敲低导致7天内通过细胞计数测量的增殖能力显著增加。研究者在PINK1敲低细胞和对照细胞中未观察到线粒体拷贝数或细胞大小的变化。接下来,使用可渗透细胞的荧光探针测量氧化应激和ROS水平。PINK1敲低细胞具有显著升高的内源性ROS水平,与胶质母细胞瘤细胞相当,而对照细胞和PINK1拯救细胞的ROS水平则显著较低。
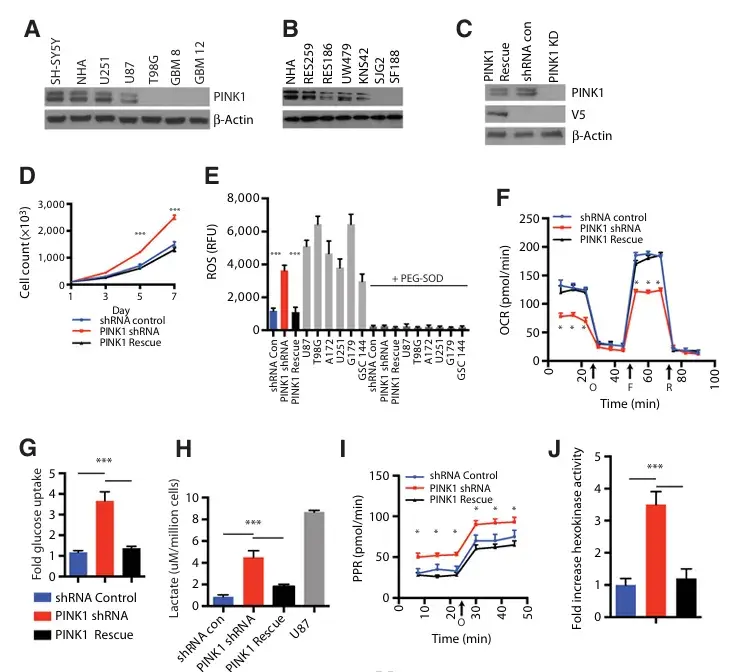
PINK1的缺失会改变星形胶质细胞的正常代谢。(A)Western印迹分析显示NHAs(正常人类星形胶质细胞)、胶质母细胞瘤细胞系和胶质母细胞瘤组织样本(GBM8和GBM12)中PINK1的表达存在差异。(B)针对儿科胶质母细胞瘤细胞系中PINK1的Western印迹分析。(C)对NHAs中通过稳定shRNA敲低PINK1的结果进行Western印迹分析,与shRNA控制的NHAs和带有V5标签的PINK1恢复NHAs进行比较。(D)对PINK1 shRNA NHAs与对照shRNA NHAs进行细胞计数分析。(E)对NHA的对照shRNA、PINK1敲低细胞和胶质母细胞瘤细胞进行了内源性ROS检测。测量工作是通过使用一种对ROS敏感的探针(1 μmol/L 氯甲基-H2DCFDA)来进行的。荧光ROS信号在加入100 U/mL PEG-SOD后被猝灭。(F)NHAs与带有PINK1敲除的NHAs在暴露于不同生物能量调节剂(如寡霉素(O)、羰基氰基-对-三氟甲氧基苯腙(FCCP/F)和罗丹明(R))后所呈现的OCR(耗氧率)比较。箭头标记化合物添加的时点。(G)通过用荧光葡萄糖类似物2-NBDG进行孵育来测量细胞中的葡萄糖摄取量。U87细胞被用作阳性对照。(H)图D中描述的细胞中的细胞外乳酸含量是通过一种NADH耦联酶反应方法进行测量的,并于第3天以490 nm的吸光度值进行测定,同时将其与细胞数量进行归一化处理。U87细胞被用作阳性对照。(I)糖酵解通量分析测量。在U87空载体对照细胞与U87 PINK1过表达细胞之间比较,测定了质子生成速率(PPR,pmol/min)。(J)总HK(己糖激酶)活性的倍数增加情况,以细胞中的随机shRNA为参照进行归一化处理
为了将探针氧化的变化归因于ROS,当实验重复并添加100 U/mL PEG-SOD时,荧光信号被有效阻断。作为荧光信号的附加对照,研究人员用氧化不敏感类似物探针重复实验,未观察到组间荧光变化,这有力支持了基于DCFH2的ROS测量是由于探针氧化的结论。用线粒体抗氧化剂mito-TEMPO处理PINK1敲低细胞可抑制ROS升高,但用NADPH氧化酶抑制剂处理时则不能,这支持了PINK1抑制线粒体ROS的来源。在PINK敲低细胞中过表达HA标记的线粒体超氧化物歧化酶2(SOD2)可抑制ROS,并完全拯救了由PINK1敲低诱导的几种表型,即细胞生长、葡萄糖摄取、乳酸产生、己糖激酶和丙酮酸激酶活性等均恢复至接近正常水平。
与对照组相比,PINK1缺失导致基础耗氧率(OCR)显著降低,最大基础糖酵解能力也显著降低,但当用寡霉素处理时,糖酵解能力反而更高。在敲低细胞中观察到己糖激酶(HK)活性增加和几种糖酵解基因的表达上调,但在对照细胞中未观察到类似变化。
PINK1缺失促进PKM2失活并稳定HIF1A
丙酮酸激酶M2亚型(PKM2)、HIF1A及其靶基因已知在有氧糖酵解过程中至关重要。多项研究表明,与非转化细胞相比,癌细胞具有上调的糖酵解水平和降低的PKM2活性,这与乳酸生成增加密切相关。在高糖酵解细胞中观察到PKM2活性降低的一种重要机制是由ROS介导的。与对照和拯救细胞相比,PINK1敲低细胞的总丙酮酸激酶活性显著降低。本研究的NHA细胞主要表达PKM2而非PKM1,这使得研究人员的测定结果对PKM2活性具有高度特异性。
![K,对盟shRNANHAs与PINK1 shRNA NHAs的丙酮酸激陶活性测定结果对比。PKM活性测定于第3天进行,并以10克细胞裂解物为参照进行归一化处理。 L,1%65DS中PKM2的免疫沉淀分析,通过标准 Westerm 方法与抗半胱氨酸砜抗体进行印迹,显示在有或无H202 (1mmolL)、氧化剂及N乙酰半胱氨酸(NAC,5mmoUL)存在的情况下PKM2上的氧化半跳氨酸残基。 M,Westem印迹结果显示,在暴露于正常氧环境和不同时间点的缺氧(氧分压为1%602)条件下,对照细胞和PINKI敲低细胞中HIF1A的稳定性均得到维持。 N,对 PINK] shRNA NHAs与对照shRNA NHAs在1%缺氧环境下的细胞计数进行对比分析的结果,P<0.05; *,P<0.01;**,P0.001](https://www.incsg.com/uploads/allimg/260509/3-260509102I2538.jpg)
(K)对乱序shRNA NHAs与PINK1 shRNA NHAs的丙酮酸激酶(PKM)活性测定结果对比。PKM活性测定于第3天进行,并以10 μg细胞裂解物为参照进行归一化处理。(L)在1% DSS中PKM2的免疫沉淀分析,通过标准Western方法与抗半胱氨酸砜抗体进行印迹,显示在有或无H2O2(1 mmol/L)、氧化剂及N-乙酰半胱氨酸(NAC, 5 mmol/L)存在的情况下PKM2上的氧化半胱氨酸残基水平。(M)Western印迹结果显示,在暴露于正常氧环境和不同时间点的缺氧(氧分压为1% O2)条件下,对照细胞和PINK1敲低细胞中HIF1A的稳定性均得到维持。(N)对PINK1 shRNA NHAs与对照shRNA NHAs在1%缺氧环境下的细胞计数进行对比分析的结果。*P < 0.05;**P < 0.01;***P < 0.001
ROS是PKM2的负调控因子,可氧化PKM2上的半胱氨酸残基,阻止其有效活性所需的四聚化过程。研究者假设PINK1敲低细胞中升高的ROS可能通过氧化半胱氨酸氨基酸来降低PKM2活性。研究者在来自PINK1敲低细胞的PKM2免疫沉淀物中检测到氧化的半胱氨酸氨基酸,但在对照组中未检测到。用过氧化氢(H2O2)处理进一步增加了PKM2中氧化半胱氨酸的水平。这种增加被强还原剂N-乙酰半胱氨酸(NAC)的添加所消除。升高的ROS水平同时也被证实可增加和稳定HIF1A信号。研究者还在常氧条件下在PINK1敲低的NHA中观察到HIF1A蛋白水平显著增加,但在对照细胞中未观察到类似现象。在1%低氧条件下处理的PINK1敲低细胞在多个时间点维持显著增加的HIF1A蛋白水平,并且在低氧期间增殖速度加快。作为验证,调节向糖酵解代谢转变的HIF1A靶基因(包括LDHA、PDK1和PKM2)在PINK1敲低细胞中均上调。当PINK1敲低细胞在常氧和低氧条件下用HIF1A siRNA处理时,细胞增殖和侵袭能力得到部分拯救。用NAC处理的PINK1敲低细胞在常氧和低氧条件下HIF1A蛋白水平均降低,导致ROS减少和细胞倍增时间延长。

- 所属栏目:神外资讯
- 如想转载“表达PINK1蛋白的胶质母细胞瘤患者生存结局更好(上)”请务必注明来源和链接。
- 网址:https://www.incsg.com/shenwaizixun/8279.html
- 更新时间:2026-05-09 10:20:11




